It’s been ages since my last post. This is because I’ve been busy doing lots of interesting physics, met a bunch of interesting physicists, maybe I’ll write something about it. For now, something I’ve been meaning to write about for a while, and for once it’s something that’s timely.
The journal Soft Matter has an issue out with a membrane biophysics theme. You can read the editorial for yourself if you have access, otherwise make do with my ropey understanding of it. Soft Matter is a relatively new journal that I think is looking really good. Their website needs work but I’ll leave that for my science 2.0 rant which is bubbling up.
So why am I interested in membranes (I’m not working on them, I’m just interested)? Well once again I’m interested in them as large system of small parts that make something amazing when they get together - ie statistical physics. So, here’s my compressed guide to membranes: please remember I’m not a biologist, I’m very new to this, only barely understand it and I tend to over simplify things.
What does the membrane do?
The cell membrane is what keeps the cell plasma and the outside separated. It has to selectively let in and out things that the cell needs or wants rid of. It also handles communications, controls electrical potential and a host of exciting things. It really is quite a complex beast.
What’s it made from?
Your basic building blocks are the lipids. We’re interested in the amphiphilic ones. They have hydrophilic heads that like water and hydrophobic tails that don’t. See the pretty MS Paint picture below.
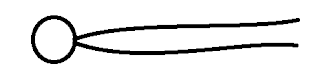
If you put a bunch of them in water then they will self assemble a
number of different ways depending on the conditions. This on its own is
really interesting. They can all go into little balls protecting the
tails from the water (micelle).
They can also form a bilayer sheet, with all the tails pointing inwards
- this is what cell membranes are made of. Picture below from the
micelle wikipedia page.
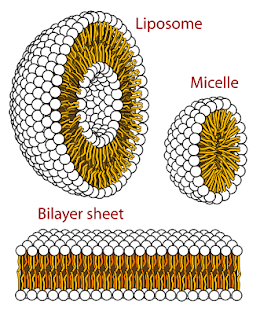
In a real cell it is of course a little more complicated. Membranes are typically made from two kinds of lipid (phospholipids and sphingolipids, the difference seems to be tail length) and on the membrane are scattered proteins that cut through the sheet that perform all manner of functions. On the outside of the membrane the heads are mostly in a two-dimensional liquid kind of organisation (more in moment). Inside the bilayer you get cholesterol floating around and the tails can be ordered or disordered - again, more on this in a moment.
Phases
Within a lipid bilayer we can have two phases that will depend on the temperature. Strictly speaking you have to be careful when you talk about a phase in a small system, but let’s not worry about that for now. At higher temperatures we have a state where all the lipid tails are disordered and wiggling around. This is the liquid-disordered phase, lipids have a very high mobility and wander around the surface. If you drop the temperature then the tails all go straight and line up. I think the heads form a crystal structure on the surface but the biologists are a little unclear on this. It’s possible the heads arrest in a gel state but I’m not sure. Either way, they stop moving around - this is the solid phase.
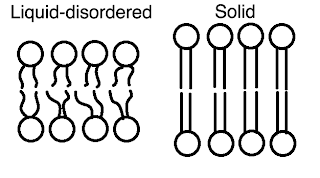 Next,
if you add cholesterol, then you can get a third phase. The so-called
liquid-ordered phase. Here our lipids tails straighten up like the solid
but there are lots of cholesterols getting into the gaps between them.
This apparently makes a phase that is ordered yet remains a liquid with
a high surface mobility. So, we have solid (SO), liquid-disordered (LD)
and liquid ordered (LO).
Next,
if you add cholesterol, then you can get a third phase. The so-called
liquid-ordered phase. Here our lipids tails straighten up like the solid
but there are lots of cholesterols getting into the gaps between them.
This apparently makes a phase that is ordered yet remains a liquid with
a high surface mobility. So, we have solid (SO), liquid-disordered (LD)
and liquid ordered (LO).
Finally we can have a mixture of species with different melting temperatures. This is what real cells are meant to be like. I have to say I find all these phases a little confusing and the terminology a little ambiguous. Anyway, stick with me, it gets more interesting from here on.
Rafts
The reason I’ve been going on about phases is because it seems to be important for the function of the cell. It is thought that on real cell membranes there exist “rafts”, small domains of liquid-ordered lipids that perform important functions such as “signaling, recruitment of specific proteins and endocytosis”. I don’t even know what endocytosis is, which is why we’ll now leave cells for something a bit simpler.
GUVs
In order to separate what’s going on it’s possible to grow simplified membranes by using giant unilamellar vesicles (GUVs). Essentially you can make a large sphere with mixtures of lipids and cholesterol but without the proteins and other stuff that’s on a real cell. And you can make them seemingly arbitrarily large.
It turns out that with the two species of lipid and the cholesterol you can get very complicated phase behaviour. Depending on the conditions you can get SO-LD coexistence, SO-LO and the most interesting, LO-LD coexistence. As someone that deals with phase coexistence on a daily basis this is very exciting. I really don’t want to get into the details so I’ll just show some cool pictures from this paper by the Keller group. If my readership ever reaches double figures I might need to think about asking for permission for these things…

Here we have LO-LD coexistence with small domains of LO. You can even get critical points!

There’s lots of things I still want to understand. I don’t even really know what the interactions are that are driving these phase transitions. There are lots of factors including curvature, surface tensions and all sorts that I haven’t thought much about yet. I think this is a really interesting area that any statistical physicist can’t fail to be excited by. It’s encouraging to see some people who appear to be able to link two such different subjects working on it.
Here are links to the main papers I’ve been learning from.
Veatch and Keller,
great paper on the GUVs.
Bagatolli and
Kumar,
very nice review in soft matter.
Semrau and
Schmidt,
another good review. Didn’t get to the end sadly.
And complete phase diagrams are in another paper by Veatch and Keller